Опишите особенности строения и функции ретикулярной формации. Функции ретикулярной формации
Наряду с быстро реагирующей на раздражители первой активизирующей системой, в которую входят проводящие пути, существует еще и неспецифическая система замедленного ответа на внешние импульсы, которая является филогенетически более древней, чем остальные структуры мозга и напоминает диффузный тип нервной системы. Эта структура называется ретикулярной формацией (РФ) и состоит из более чем 100 ядер, соединенных между собой. РФ простирается от ядер таламуса и субталамуса до промежуточной зоны спинного мозга верхних шейных сегментов.
Первые описания РФ были сделаны немецкими морфологами: в 1861 г. К. Рейхертом и в 1863 г. О. Дейтерсом, который и ввел термин РФ; большой вклад в ее изучение внес В.М. Бехтерев.
Нейроны, составляющие РФ разнообразны по величине, строению и функции; имеют обширно разветвленное дендритное дерево и длинные аксоны; их отростки густо переплетаются, напоминая сеть (лат. reticulum - сеточка, formatio - образование).
Свойства ретикулярных нейронов :
1. Мультипликация (умножение импульса) и амплификация (получение конечного большого результата) – осуществляется благодаря сложному переплетению отростков нейронов. Поступающий импульс многократно умножается, что в восходящем направлении дает ощущение даже небольших стимулов, а в нисходящем направлении (ретикулоспинальные пути) позволяет вовлекать в ответную реакцию множество структур НС.
2. Генерирование импульсов . Д. Моруцци доказал, что большинство нейронов РФ постоянно генерирует нервные разряды с частотой порядка 5-10 в секунду. Различные афферентные стимулы суммируются с этой фоновой активностью ретикулярных нейронов, вызывая в одних из них ее учащение, а в других торможение.
3. Полисенсорность . Почти все нейроны РФ способны отвечать на на раздражения самых различных рецепторов. Тем не менее, часть из них реагируют на кожные раздражения и на свет, другие на звуковые и кожные раздражения и т. д. Таким образом, полного смешивания афферентных сигналов в ретикулярных нейронах не происходит; в их связях есть частичная внутренняя дифференциация.
4. Чувствительность к гуморальным факторам и, особенно, к фармпрепаратам. Особенно активными оказываются соединения барбитуровой кислоты, которая даже в небольших концентрациях полностью прекращают активность ретикулярных нейронов, при этом не действуя на спинальные нейроны или на нейроны коры больших полушарий.
В целом же, РФ характеризуется разлитыми рецептивными полями, длительным латентным периодом ответа на периферическую стимуляцию и слабой воспроизводимостью реакции.
Классификация :
Существует топографическая и функциональная класификация РФ.
I. Топографически вся ретикулярная формация может быть подразделена на каудальный и ростральный отделы.
1. Ростральные ядра (ядра среднего мозга и верхней части моста, связанные с промежуточным мозгом) – отвечают за состояние возбуждения, бодрствования, настороженности. Ростральные ядра оказывают локальное влияние на определенные зоны коры больших полушарий. Поражение этого отдела вызывает сонливость.
2. Каудальные ядра (моста и промежуточного мозга, связаны с ядрами черепных нервов и спинным мозгом) – выполняют моторную, рефлекторную и вегетативную функции. Некоторые ядра в процессе эволюции получили специализацию - вазомоторный центр (депрессорные и прессорные зоны), дыхательный центр (экспираторный и инспираторный), рвотный центр. Каудальный отдел РФ оказывает более разлитое, генерализованное влияние на обширные зоны головного мозга. Поражение этого отдела вызывает бессонницу.
Если рассматривать ядра РФ каждого отдела мозга, то РФ таламуса образуют капсулу латерально вокруг зрительных бугров. Они получают импульсы от коры и дорзальных ядер таламуса. Функция ретикулярных ядер таламуса – фильтрация сигналов, идущих через таламус к коре полушарий; проекция их на другие ядра таламуса. В целом, они влияют на всю поступающую сенсорную и когнитивную информацию.
Ядра РФ среднего мозга включают покрышечные ядра: nuclei tegmentalis dorsalis et ventralis , nucleus cuneiformis . Они получают импульсы через fasciculus mammillo-tegmentalis (Gudden) , который является частью мамиллоталямического пути.
РФ моста, образованная околосрединными (парамедианными) ядрами, не имеет четких границ. Эти ядра участвуют в скоординированном движении глаз, фиксированном взгляде и саккадном движении глаз (быстрые синхронные движения глаз). РФ моста лежит кпереди и латерально от медиального продольного пучка, получает импульсы по нервным волокнам от верхних бугорков четверохолмия через предорзальные нервные волокна и от передних зрительных полей посредством лобномостовых связей.
Латеральную РФ образуют преимущественно ядра РФ продолговатого мозга. Эта структура имеет множество ганглиев, вставочных нейронов вокруг черепных нервов, которые служат для модулирования связанных с ними рефлексов и функций.
II. Функционально ядра РФ подразделяются на вертикальные образования:
1. Срединный столб (ядра шва) – узкий парный столб клеток вдоль срединной линии ствола мозга. Простираются от продолговатого до среднего мозга. Дорзальные ядра шва синтезируют серотонин.
2. Медиальный столб (голубоватое пятно) – относится к РФ. Клетки голубоватого пятна синтезируют норэпинефрин, аксоны идут к областям коры, ответственным за возбуждение (бодрствование).
3. Латеральный столб (серое вещество вокруг Сильвиева водопровода) – (входит в состав лимбической системы) – клетки имеют опиоидные рецепторы, что способствует эффекту обезболивания.
Функция РФ:
1. Регуляция сознания путем изменения активности корковых нейронов, участие в цикле сон /бодрствование, возбуждение, внимание, обучаемость - когнитивные функции
2. Обеспечение эмоциональной окраски сенсорным стимулам (ретикулолимбические связи)
3. Участие в жизненно важных вегетативных реакциях (сосудодвигательный, дыхательный, кашлевой, рвотный центры)
4. Реакция на боль – РФ проводит болевые импульсы к коре и образует нисходящие аналгезирующие пути (влияет на спинной мозг, частично блокируя передачу болевых импульсов от спинного мозга к коре)
5. Привыкание – это процесс, в котором мозг учится игнорировать незначительные повторяющиеся стимулы извне, отдавая предпочтение новым стимулам. Пример – способность спать в переполненном шумном транспорте, сохраняя способность проснуться на сигнал автомобиля или крик ребенка
6. Соматодвигательный контроль – обеспечивается ретикулоспинальными путями. Эти пути отвечают за мышечный тонус, равновесие, положение тела в пространстве, особенно при его движении
7. Формирование интегрированных реакций организма на стимулы, например, сочетанная работа речедвигательного аппарата, обшая двигательная активность.
Связи РФ
Аксоны РФ связывают практически все структуры мозга между собой. РФ морфологически и функционально связана со спинным мозгом, мозжечком, лимбической системой и корой больших полушарий головного мозга.
Часть аксонов РФ имеет нисходящее направление и образует ретикулоспинальные пути, а часть восходящее (спиноретикулярные пути). Возможна также циркуляция импульсов по замкнутым нейронным цепям. Таким образом, существует постоянный уровень возбуждения нейронов РФ, вследствие чего обеспечиваются тонус и определённая степень готовности к деятельности различных отделов центральной нервной системы. Степень возбуждения РФ регулируется корой больших полушарий головного мозга.
1. Спиноретикулярные (спиноретикулокортикальные) пути (восходящая активизирующая ретикулярная система) - получают импульсы от аксонов восходящих (сенсорных) путей общей и специальной чувствительности. Соматовисцеральные волокна идут в составе спиноретикулярного тракта (переднебоковой канатик), а также в составе проприоспинальных путей и соответствующих путей от ядра спинального тройничного тракта. К ретикулярной формации приходят также пути от всех других афферентных черепномозговых нервов, т.е. практически от всех органов чувств. Дополнительная афферентация поступает от многих других отделов головного мозга - от моторных областей коры и сенсорных областей коры, мозжечка, базальных ядер, красного ядра, от таламуса и гипоталамуса. Эта часть РФ отвечает за процессы возбуждения, внимания, бодрствования, обеспечивает эмоциональные реакции, важные при познавательном процессе. Поражения, опухоли этой части РФ вызывают снижение уровня сознания, активности психической деятельности, в частности когнитивных функций, двигательной активности, синдром хронической усталости. Возможны сонливость, проявления оглушенности, общей и речевой гипокинезии, акинетического мутизма, сопора, в тяжелых случаях - кома.
2. Ретикулоспинальные пути (нисходящие ретикулярные связи) – могут оказывать как стимулирующее влияние (отвечают за мышечный тонус, вегетативные функции, активирует восходящую РФ), так и угнетающее (способствуют плавности и точности произвольных движений, регулирует мышечный тонус, положение тела в пространстве, вегетативные функции, рефлексы). Обеспечиваются множеством эфферентных связей - нисходящие к спинному мозгу и восходящие через неспецифические таламические ядра к коре головного мозга, гипоталамусу и лимбической системе. Большинство нейронов образует синапсы с двумя - тремя дендритами разного происхождения, такая полисенсорная конвергенция характерна для нейронов ретикулярной формации.
3. Ретикуло-ретикулярные связи .
Ретикулярная формация ствола мозга представляет собой комплекс нейронов, имеющих обширные связи с разными нервными центрами, друг с другом и корой полушарий. Она пролегает в ростральном направлении к таламусу. Рассмотрим далее ее особенности.
Функции ретикулярной формации
В задачи комплекса входит обработка сенсорной информации. Кроме этого, ретикулярная формация обеспечивает активизирующее воздействие на кору, осуществляя контроль деятельности спинного мозга. За счет этого регулируются тонус скелетных мышц, работа вегетативной и половой систем человека.
Механизм действия
Впервые он был выявлен Р. Гранитом. Ученый установил, что может влиять на активность γ-мотонейронов. Вследствие этого γ-эфференты (их аксоны) провоцируют сокращение веретен мускулатуры и, соответственно, повышение афферентной импульсации мышечных рецепторов. Поступающие в спинной мозг сигналы провоцируют возбуждение α-мотонейронов. Это и обуславливает тонус мускулатуры. Было установлено, что в реализации этой функции участвуют нейроны формации моста и продолговатого мозга. Их поведение диаметрально противоположно. Последние провоцируют активацию α-мотонейронов в мышцах-сгибателях и, соответственно, тормозят их в разгибателях. Нейроны моста действуют наоборот. Ретикулярная формация связана с мозжечком и корой, от которой поступает информация. Это позволяет сделать вывод, что она выступает в качестве коллектора неспецифического сенсорного потока, который, возможно, участвует в регуляции активности мускулатуры. Однако в настоящее время еще не выяснена необходимость формации, дублирующей задачи нейронов в красном и вестибулярных ядрах.

Структура
Ретикулярная формация образуется рассеянными клетками. Некоторые из них считаются жизненно важными образованиями. В частности, можно выделить центры:
- Дыхательный и сосудодвигательный. Они располагаются в продолговатом мозге.
- Координации взора. Он находится в среднем мозге.
- Голода, насыщения и терморегуляции. Они располагаются в промежуточном мозге.
В качестве ключевого тракта выступает ретикулоспинальный. Он проходит к нейронам в двигательных ядрах передних спинномозговых рогов и черепных нервов по стволу и к вставочным элементам нервной вегетативной системы. От них пролегают таламо-корковые волокна. Они обеспечивают активацию коры, которая необходима для восприятия специфических раздражителей. Эти таламо-корковые волокна заканчиваются во всех корковых слоях.

Научные наблюдения
В ходе исследований было выявлено, что ретикулярная формация обладает активизирующим воздействием на кору. Этот нейронный комплекс выступает в качестве своеобразного "энергетического центра". Без него нервные клетки коры, разные ее отделы, а также весь мозг в целом не смогут выполнять все свои многообразные сложные задачи. Комплекс нейронов непосредственно участвует в процессе регулирования сна и бодрствования. Результаты экспериментов позволили объяснить некоторые наблюдения хирургов. Так, в процессе операций на мозге могут быть сделаны разрезы в коре полушарий, удалена часть ткани. При этом пациент сознания не потеряет. Однако, если скальпелем будет задета , человек впадет в глубокий сон.
Специфика работы
Сегодня достаточно хорошо изучены специфические нервные каналы, по которым от органов чувств передается информация в мозг. Именно так кора узнает о характере раздражителя, действующего на организм. В соответствии с этим она посылает разные импульсы к системам и органам. Исследования показали, что от всех волокон, направленных от периферии к коре, отходят ответвления. Они заканчиваются на поверхности клеток формации. Внешнее раздражение любого характера оказывает на нее возбуждающее действие. В этот момент происходит своего рода "зарядка энергией". Выступая как мозговой центр, формация определяет степень работоспособности коры. Активизируя все отделы, она обеспечивает точный синтез и анализ многообразия информации, которая поступает в кору из внешнего мира.

Реакция на вещества организма
Ретикулярная формация чувствительна не только к нервным сигналам, но и к растворенным в крови соединениям. В частности, речь о сахаре, гормонах, углекислоте, кислороде. Особое значение среди этих веществ имеет адреналин. При эмоциональном перенапряжении - при гневе, страхе, состоянии аффекта, ярости - отмечается продолжительное возбуждение формации. Его поддерживает адреналин, усиленно выделяющийся в кровь. Активность комплекса во многом определяют и другие химические соединения. В первую очередь это углекислый газ и кислород. К примеру, если у человека во сне затруднено дыхание, то СО 2 начинает накапливаться в крови. Углекислота активизирует ретикулярную формацию, вследствие чего человек просыпается.

Заключение
Клинические исследования и экспериментальные данные, полученные в физиологических лабораториях, показали, что ретикулярная формация прямо связана с возникновением эмоций. Итоги изучения ее строения и задач, которые она реализует, широко применяются в психо- и нейрофармакологии. Было установлено, что вялость, апатия, сонливость или раздражительность, бессонница могут обуславливаться расстройством в работе ретикулярной формации. Этот нейронный комплекс также выполняет определенную роль в процессе возникновения многих патологий ЦНС.
Билет 15
1. Формы (фрагменты) афферентного синтеза: Доминирующая мотивация; Обстановочная афферентация; Пусковая афферентация. Роль ретикулярной формации.
2. Быстрые и медленные мышечные волокна.
Вопрос 1
АФФЕРЕНТНЫЙ СИНТЕЗ - (соединение, составление) - процесс сопоставления, отбора и синтеза многочисленных и разных по функциональному значению афферентаций, вызванных различными воздействиями на организм, происходящий в ц. н. с., на основе к-рого формируется цель действия.
А. с. согласно теории функциональной системы Анохина - первая, универсальная, стадия любого целенаправленного поведенческого акта (см. Функциональные системы).
А. с. включает обработку 4 основных видов афферентных возбуждений.
1. Мотивационное возбуждение отражает доминирующую потребность организма, к-рая возникает под влиянием метаболических, гормональных, а у человека - и социальных факторов Мотивация играет решающую роль в формировании цели действия. Специфически повышая реактивность корковых нейронов с помощью ориентировочно-исследовательской реакции , мотивационное возбуждение способствует обработке и активному отбору сенсорной информации, необходимой для построения целенаправленного поведения.
2. Обстановочная афферентация представляет собой воздействие на организм всей совокупности внешних факторов, составляющих конкретную обстановку, на фоне к-рой развертывается приспособительная деятельность. Обстановочная афферентация формируется не только постоянными компонентами обстановки, но и рядом последовательных афферентных воздействий на организм. Характерная особенность обстановочной афферентации состоит в том, что она придает специфику будущей поведенческой реакции, обеспечивая ее приспособительное значение только в данной обстановке.
Наиболее отчетливо роль обстановочной афферентации проявляется в опытах с условными рефлексами. В этих случаях на один и тот же условный раздражитель животное отвечает условной оборонительной реакцией в одной экспериментальной камере и условной пищевой - в другой (или в одной и той же экспериментальной камере утром животное отвечает пищевой реакцией, а вечером - оборонительной).
На стадии афферентного синтеза решаются вопросы “что делать?”, “как делать?”, “когда делать?”.
Пусковая афферентация
Представляет собой специальный раздражитель, собственно запускающий поведенческую реакцию. Значение пускового раздражителя состоит в том, что он призван обозначить момент начала поведенческой реакции.
Целенаправленное поведение может начинаться и без явного пускового стимула. Примерами таких реакций являются регулярно совершающиеся физиологические отправления (еда, сон, дефекация, мочеиспускание и др.), приуроченные к определенным периодам суток.
Афферентный синтез осуществляется на основе следующих нейрофизиологических механизмов:
1) механизмы восходящих активирующих влияний подкорковых образований на кору головного мозга. Это прежде всего, активирующие влияния гипоталамуса к лобным отделам коры, через передние ядра таламуса, что отражает мотивационные возбуждения. Аналогичным образом воздействуют другие лимбические системы. Вторыми по активирующему значению являются ретикулярные структуры среднего мозга и моста, которые обеспечивают соответствующий уровень бодрствования.
2) механизмы конвергенции возбуждений различного качества на нейронах коры и подкорковых структурах мозга. В частности, мультисенсорная конвергенция от поверхностей (зрительная, тактильная, слуховая, температурная и др.); мультибиологическая конвергенция, связанная с определенными состояниями (голод, боль и т.п.) и др.;
3) интеграцию мотивационных, обстановочных и пусковых афферентаций на нейронах коры мозга;
4) механизмы формирования доминанты, за счет которых подавляется текущая деятельность и удерживается вновь сформированная поведенческая реакция.
Роль ретикулярной формации
Ретикулярная формация характеризуется относительно малой возбудимостью. Эффекты ее раздражения появляются через большой латентный период, она медленно реагирует и остается активной в течение продолжительного времени после прекращения раздражения (длительное последействие). Ретикулярная формация облегчает или подавляет фазные движения и напряжение скелетных мышц, вызываемые мотонейронами спинного мозга, а также движения, вызванные с коры больших полушарий. Ретикулярная формация среднего и промежуточного мозга облегчает рефлекторные движения животных, раздражение промежуточного мозга тормозит двигательные рефлексы спинного мозга.
Боковые отделы ретикулярной формации варолиева моста и среднего мозга облегчают, а средние ее отделы в продолговатом мозге тормозят двигательные рефлексы. Облегчение и торможение зависят также от интенсивности и продолжительности раздражения ретикулярной формации. По гамма-нейронам она регулирует функции мышечных веретен, следовательно, обратную информацию из скелетных мышц. Она изменяет также возбудимость восходящих афферентных путей спинного мозга, что может снизить или прекратить постсинаптическое торможение. Тонические влияния ретикулярной формации вызывают ВПСП или ТПСП в мотонейронах спинного мозга. Она изменяет также передачу импульсов в мозговом стволе и одновременно с влиянием на скелетную мускулатуру вызывает сосудодвигательные, дыхательные, зрачковые и другие реакции.
Ретикулярная формация оказывает адаптационно-трофическое влияние на кору больших полушарий, подкорковые образования промежуточного мозга, мозжечок и спинной мозг. Существуют взаимные влияния этих отделов нервной системы, как возбуждающие, так и тормозящие. Она участвует в физиологических процессах сна и пробуждения, а также в эмоциях, в реакции напряжения («стресс») и др. Раздражение ретикулярной формации вызывает пробуждение спящих животных, а ее разрушение и выключение - глубокий сон у бодрствующих животных. Изучены взаимные влияния ретикулярной формации и коры больших полушарий.Установлено участие ретикулярной формации в образовании и протекании условных рефлексов
По симпатическим волокнам ретикулярная формация регулирует возбудимость и работоспособность скелетной мускулатуры, функциональное состояние нервной системы и органов чувств, оказывая на них адаптационно-трофическое влияние. Регуляция рефлексов позы и двигательных рефлексов, перемещающих тело, осуществляется по эфферентным гамма-волокнам иннервирующим проприоцепторы.
Ретикулярная формация регулирует вегетативные функции, деятельность внутренних органов. Она влияет на образование гормонов в гипофизе и других железах внутренней секреции и в ней концентрируются гормоны и медиаторы.
Афферентные волокна поступают в нее по симпатическим и блуждающим нервам. Частьклеток ретикулярной формации среднего мозга и варолнева моста возбуждается адреналином и норадреналином (адренореактивные системы) а другая часть, расположенная в промежуточном мозге, несколько выше среднего мозга, возбуждается ацетилхолином и его производными (холинореактивные системы). Адренореактивные системы среднего мозга и варолиева моста облегчают наступление двигательных рефлексов, а адренореактивные системы продолговатого мозга тормозят спинномозговые рефлексы. Адреналин возбуждает и холинореактивные системы. Предполагается, что действие ацетилхолина и его производных менее ограничено, чем действие адреналина, и охватывает многие области головного мозга. Действие ацетилхолина на ретикулярную формацию противоположно его периферическому влиянию на внутренние органы. Ретикулярную формацию среднего и продолговатого мозга возбуждает углекислота.
Гормоны и медиаторы действуют на функцию больших полушарий как непосредственно, так и посредством ретикулярной формации. Таким образом, ретикулярная формация мозгового ствола - подкорковый центр вегетативной нервной системы.
Вопрос 2 .
Сложное строение человеческого мозга открывает тайны нашего поведения, объясняет законы мыслительной деятельности, протекание эмоций и чувств. Каждое полушарие головного мозга отвечает за свои специфические функции и задачи (например, известно, что правое отвечает за логику, а левое - за воображение и фантазию), но имеются также структуры, которые обеспечивают единую и согласованную работу всей центральной нервной системы. Одной из таких структур является ретикулярная формация.
Общие сведения
Ретикулярная формация - это участок стволового отдела мозга, представленный разветвленной сетью нервных клеток и ядер, соединяющих между собой различные отделы головного мозга. В отличие от других структур, - например, таламуса, гипоталамуса, мозжечка, - которые имеют некую цельную форму (ядра, железы), ретикулярная формация не представлена единым морфологическим образованием, а является «сетью» (от латинского reticulum — сеть) дендритов и аксонов, которые с разной степенью плотности проникают между отделами и структурами головного мозга, объединяя их между собой и обеспечивая их совместную деятельность.
Метафорически выражаясь: если наш мозг представить в виде некоего изделия, скажем, рубашки, то ретикулярная формация - это нити, которыми рубашка сшита. Ретикулярная формация пронизывает структуры продолговатого, среднего мозга и моста, имеет непосредственные связи с мозжечком, спинным мозгом, таламусом и опосредованные - с вышележащими отделами: гипоталамусом, зрительными ядрами и корой.
Как устроена
В состав ретикулярной формации входит огромное количество нейронов с разветвленными дендритами и длинными аксонами, за счет чего становится возможным передавать нервные импульсы в различные отделы головного и спинного мозга. При этом можно выделить две наиболее крупные группы нейронных скоплений:
- Ретикулотегментальное ядро, нейроны которого получают сигналы от вышележащих отделов ГМ (четверохолмие, таламус) и передают их далее в структуры мозжечка, регулируя тем самым некоторые жизненно важные двигательные функции: координация взгляда, движения глаз.
- Латеральное ядро, нейроны которого восходят от структур спинного мозга и вестибулярных ядер и обеспечивают информирование коры ГМ о положении тела в пространстве, участвуют в регуляции дыхания и сосудистой иннервации.
- Кроме того, в состав ретикулярной формации входят нейроны, которые принимают важное участие в работе центров терморегуляции, насыщения и голода.

Основные функции
Основное предназначение ретикулярной формации состоит в сенсорном анализе многочисленных сигналов, поступающих от различных отделов ГМ.
За счет тесных связей со спинным мозгом она также принимает самое активное участие в моторной регуляции, начиная от глотательного рефлекса и заканчивая сложными двигательными операциями. Кроме того, ретикулярная формация осуществляет активирующее воздействие на весь ГМ в целом, участвуя в регуляции циклов сна и бодрствования.
В общем виде функции ретикулярной формации сводятся к следующим:
- Регуляция скелетной мускулатуры (участвует в управлении движениями тела) и вегетативных функций (дыхание, чихание, кровообращение и т.п.).
- Контроль процессов сна и бодрствования (за счет оказания активирующего и тормозящего воздействия на кору больших полушарий).
- Активизирующая функция (проявляется в том, что ретикулярная формация обеспечивает постоянное тоническое возбуждение коры ГМ, за счет чего становится возможным поддержание внимания, сознания и протекание мыслительных процессов.)
- Обработка сигналов, поступающих из внешней и внутренней среды.
Отличительные особенности работы ретикулярной формации связаны, прежде всего, с определенными свойствами ее :
Нервные клетки, входящие в состав ретикулярной формации, обладают повышенной способностью к тоническому возбуждению. Это означает, что бо́льшая часть нейронов находятся в постоянном возбуждении и генерирует нервные импульсы, которые передаются в вышележащие отделы ГМ. Такая тоническая активность обусловлена рядом факторов:
- Проходимость большого количества сигналов через структуры ретикулярной формации. Приведем простую аналогию: представьте себе рояль или какой-либо другой струнный инструмент. Понятно, что когда мы напрямую касаемся струн, они начинают колебаться и издавать звук. То же самое происходит и с нервными клетками, когда к ним приходят сигналы от других нейронов.Однако представим далее, что мы не касаемся непосредственно струн инструмента, но, скажем, скачем рядом с ним, сильно стуча ногами о пол. Звука инструмента мы, может быть, и не услышим, но едва заметное колебание струн все же будет иметь место. То же самое происходит и с нейронами ретикулярной формации. Поскольку через нее постоянно проходят какие-то сигналы (как афферентные, так и эфферентные) от различных структур ЦНС, то это и создает постоянное тоническое возбуждение нейронов ретикулярной формации, за счет того, что она находится в эпицентре постоянного нервно-импульсного обмена.
- Повышенная чувствительность нейронов к химическим веществам (гормоны, медикаменты, психотропные вещества). Чашка кофе, выпитая утром, «включает» именно структуры ретикулярной формации и за счет длительного сохранения возбуждения в ее нейронах поддерживает нас в активном состоянии.
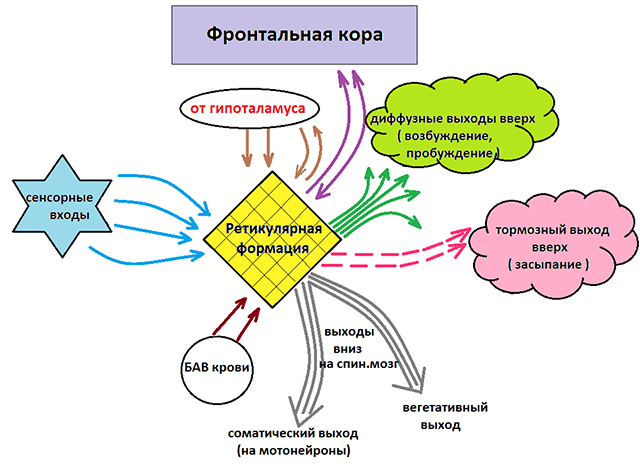
Нисходящее и восходящее влияние РФ
Как уже отмечалось, ретикулярная формация оказывает возбуждающее и тормозящее действие на различные отделы ГМ. При этом можно выделить два отдела, которые специализируются на передаче возбуждения в те или иные структуры мозга.
Нисходящий отдел: представлен вегетативными и двигательными центрами и оказывает нисходящее влияние на отделы спинного мозга. Соответствующие нейронные скопления регулируют деятельность дыхательного, сосудодвигательного, слюноотделительного центров, а также центров, отвечающих за построение простых и сложных двигательных реакций. Это указывает на решающую роль именно центральной нервной системы в регуляции даже элементарных безусловных рефлексов. Стимуляция нисходящего отдела приводит к затормаживанию спинальных центров и вызывает в естественной среде состояние глубоко сна (спать «без задних ног»). Тот же эффект можно вызвать и искусственным путем, например, вводя человека в состояние транса или наркоза.
Восходящий отдел: представлен нервными волокнами, соединяющими структуры ретикулярной формации с вышележащими отделами: таламусом, гипоталамусом, мозжечком и корой. Восходящее влияние оказывает стимулирующее действие на кортикальные структуры и обеспечивает активное состояние сознания. Восходящее влияние не прекращается, даже когда мы спим. Если бы наш мозг мог полностью «отключаться», то каждое пробуждение было бы сродни рождению: кто я? Где я? Как я сюда попал? Однако за счет работы ретикулярных структур у нас сохраняется возможность всегда возвращаться в то исходное состояние сознания, в котором мы находились до момента сна. Кроме того, и во время ночного отдыха у нас остается способность реагировать на некоторые жизненно важные раздражители, т.е. мы, как правило, не спим «мертвым сном» и можем проснуться, если рядом зашевелился и заплакал ребенок, что-то громко упало и т.п.
Проявление повреждения структур
Ретикулярная формация играет значимую роль в интегративной деятельности всего головного мозга. За счет выполнения функции ведущего проводника всех видов нервных импульсов во все отделы ЦНС, ретикулярная формация находится в постоянной работе. Чрезмерные умственные и эмоциональные перегрузки вредны для мозга в целом и для ретикулярной формации, в частности. К счастью, своевременный прием седативных препаратов может (за счет повышенной восприимчивости нейронов к химическому воздействию) быстро исправить ситуацию и нормализовать состояние.
Однако возможны и менее благоприятные исходы. Повреждение возможно в результате черепно-мозговых травм, онкологических заболеваний головного мозга, инфекционных поражений.
Основным проявлением неблагополучия является потеря сознания.
Нарушение восходящих связей обнаруживает себя в состоянии апатии, слабости, повышенной сонливости, двигательной растороможенности, нарушениях ночного сна. Нередки сопутствующие вегетативные расстройства.
РЕТИКУЛЯРНАЯ ФОРМАЦИЯ [formatio reticularis (PNA, JNA, BNA); син.: ретикулярная субстанция, сетевидное образование ] - совокупность структур, расположенных в центральных отделах спинного мозга и мозгового ствола. Для Ретикулярной Формации характерно наличие большого количества нервных волокон, проходящих в самых различных направлениях, в связи с чем данное образование под микроскопом напоминает сеточку. Это послужило основанием для Дейтерса (О. F. С. Deiters) назвать его сетчатым образованием.
Морфология
В конце 19 - начале 20 в. были описаны наружный отдел Ретикулярной Формации продолговатого мозга, состоящий преимущественно из серого вещества (formatio reticularis grisea), внутренний отдел, состоящий преимущественно из белого вещества (formatio reticularis alba), латеральный отдел (substantia reticularis lateralis), а также отдельные ядра Ретикулярной Формации. Первое систематическое описание ядер Ретикулярной Формации мозгового ствола было дано в 1909 г. Якобсоном (H. Jacobsohn).
В атласе Мессена и Ольшевского (Meessen, J. Olszewski, 1949) описано 32 ядра ромбовидного мозга кролика. В своей работе 1954 года Ольшевский описывает 22 ядра Р. ф. продолговатого мозга, моста и среднего мозга у человека.
Изучение строения отдельных ядер Ретикулярной Формации и составляющих их нейронов, а также связей Ретикулярной Формации послужило основанием для разделения Р. ф. на зоны, части или колонки.
Сравнительно-анатомическое, морфологическое и онтогенетическое изучение Р. ф. позволило В. В. Амунцу (1966, 1976, 1980) сгруппировать ядра Р. ф. в 3 отдела: наружный, внутренний и средний - и выделить переходные зоны между ретикулярными и специфическими, а также между различными ретикулярными структурами.
Характеристика нейронного строения Р. ф. была сделана Шайбелем и Шайбелем (A. B.Scheibel, М. Е. Scheibel, 1962), Г. П. Жуковой (1977), Т. А. Леонтович (1978), Манненом (H. Mannen, 1966, 1975), Вальверде (F. Valverde, 1961), Н. С.Косицыным (1976) и др.
Нервные связи Р. ф. изучались преимущественно методами дегенерации. Волокна Р. ф. делят на ра-диарные, направленные вентрально, волокна концентрические, перекрещивающиеся в области средней линии, и волокна продольные, сгруппированные в пучки. Эти пучки составляют как афферентные, так и эфферентные пути. Данные о связях Р.ф. обобщены в работах Бродала (1957), Росси и Цанкетти (G. F. Rossi, A. Zanchetti, 1957).
Физиология
В области мозгового ствола имеются анатомические образования, возбуждение к-рых оказывает генерализованное тонизирующее влияние на передние отделы головного мозга (см.). Эта группа анатомических образований получила название восходящей активирующей системы ретикулярной формации (рис. 1). Ей отводится важная роль в поддержании бодрствующего состояния, а также в механизмах формирования целостных, и в частности условнорефлекторных реакций организма. Наряду с восходящей активирующей системой выделяют также нисходящие рети-кулоспинальные системы, оказывающие контролирующее влияние на рефлекторную деятельность спинного мозга. Активность как восходящих, так и нисходящих систем поддерживается непрерывным притоком афферентных импульсов, поступающих в Р. ф. по коллатеральным волокнам от сенсорных проводящих путей. Важную роль в поддержании активности ретикулярных механизмов играют гуморальные раздражители, по отношению к к-рым Р. ф. обладает высокой чувствительностью, что обеспечивает ее участие в регуляции ряда вегетативных функций. Наряду с этим Р. ф. является местом избирательного действия многих фармакологических средств, что находит широкое применение при лечении ряда заболеваний ц. н. с., а также обусловливает новый подход к изучению таких важнейших проблем медицины, как, напр., проблема боли и обезболивания. В области Р. ф. происходит обширное пространственное совпадение и взаимодействие импульсов, поступающих от различных периферических рецепторных образований, с возбуждениями, идущими от мозжечка и коры больших полушарий. Благодаря большому количеству кортико-ретикулярных связей кора головного мозга оказывает контролирующее влияние на деятельность ретикулярных механизмов, регулируя уровень их активности.
Ретикулоспинальные соотношения. Влияние мозгового ствола на двигательную активность спинного мозга было впервые продемонстрировано И. М. Сеченовым в 1862 г. (см. Сеченовское торможение). Им было показано, что при раздражении мозгового ствола может наблюдаться как торможение, так и облегчение рефлекторных реакций. Однако механизм и структуры, опосредующие эти влияния, оставались невыясненными. В 1944 г. X. Мегун, применяя прямое электрическое раздражение мозгового ствола, показал, что раздражение определенных участков бульбарной Р. ф. приводит к полной остановке движений, вызываемых как рефлекторно, так и раздражением двигательных участков коры головного мозга. Это торможение было общим и распространялось на все группы мышц независимо от их топографических соотношений и физиологической функции. X. Мегун высказал предположение, что контролирующее влияние Р. ф. осуществляется на уровне спинного мозга, а не на уровне коры головного мозга. Это предположение было подтверждено им в опытах на децереб-рированных животных.
Относительно механизма ретику-лоспинальных влияний нет единого мнения. В то время как одни исследователи считают, что Р. ф. может оказывать влияние непосредственно на мотонейроны спинного мозга, другие предполагают, что эти влияния передаются мотонейронам через какие-то промежуточные нейроны, роль к-рых могут играть вставочные нейроны, участвующие в замыкании сегментарных спинномозговых рефлекторных дуг.
Ретикулокортикальные соотношения и проблема восходящего активирующего влияния ретикулярных механизмов на кору головного мозга. Известен факт, что основными симптомами при поражении определенных участков мозгового ствола являются общее снижение активности больного, адинамия, сонливость. Эксперименты показали, что подобные явления могут развиваться при разрушении у животного подкорково-стволовых отделов мозга. Эти данные послужили для ряда Р1сследователей основанием полагать, что в области мозгового ствола находятся центры, ведающие общей активностью организма, центры сна и бодрствования (см. Центры нервной системы). Такое предположение было обосновано тем, что прямое раздражение определенных отделов мозгового ствола могло вызвать сон у подопытного животного либо вывести его из этого состояния. Однако подлинный прогресс в этой проблеме стал возможен лишь после того, как для исследования мозга стали все более широко применять элек-трофизиологические методы, в частности электроэнцефалографию (см.). Исследования В. В. Правдич-Неминского (1925) показали, что внешние раздражения вызывают характерные изменения ЭЭГ, заключающиеся в смене медленных высокоамплитудных и низкочастотных колебаний, характерных для состояния покоя, быстрыми низкоамплитудными и высокочастотными колебаниями. Такие изменения ЭЭГ наблюдаются у человека или животного при переходе от состояния сна к бодрствованию. В связи с этим подобный тип реакции получил название «реакция активации ЭЭГ», или «реакция пробуждения».
Развитие экспериментальных методов исследования позволило разработать способы тонкого раздражения и разрушения отдельных подкорковых структур с помощью электродов, вводимых с большой точностью в нужные для экспериментатора пункты (см. Стереотаксический метод). Это позволило Дж. Мо -руцци и X. Мегуну в 1949 г. подойти к решению вопроса о тохМ, какие структуры мозга ответственны за возникновение «реакции пробуждения». Для решения этой задачи Дж. Моруцци и X. Мегун провели серию экспериментов, в ходе к-рых установили, что при раздражении у животных определенных пунктов мозгового ствола наблюдается смена медленных синхронных высоковольтных колебаний, характерных для сна, на низкоамплитудную высокочастотную активность. Эти изменения ЭЭГ были диффузными, т. е. наблюдались по всей коре головного мозга, но лучше были выражены в одноименном по отношению к участку раздражения полушарии. Изменения электрической активности коры больших полунга-рий сопровождались внешними признаками пробуждения.
Дальнейшими исследованиями показано, что подобные явления можно наблюдать при раздражении различных отделов Р. ф. мозгового ствола - начиная от продолговатого и кончая промежуточным мозгом. В области продолговатого мозга (см.) возбудимая зона совпадает с теми участками Р.ф., к-рые, по данным X. Мегуна и Райнса (R.Rhines), оказывают и нисходящее влияние на активность спинного мозга. На уровне моста (см. Мост головного мозга) и среднего мозга (см.) эта зона располагается в области покрышки, а на уровне промежуточного мозга (см.) захватывает субталамическое ядро и задний гипоталамус (см.), доходя до медиальных таламических ядер. Эти структуры мозгового ствола и составляют восходящую активирующую ретикулярную систему, анатомическим субстратом к-рой является ряд восходящих короткоаксонных ретикулярных путей. X. Мегун и Дж. Моруцци пришли к выводу, что наблюдавшиеся ими изменения ЭЭГ не являлись результатом антидромного проведения импульсов к коре больших полушарий по известным кортико-фугальным путям. Эти изменения нельзя было объяснить и проведением возбуждения к коре по известным классическим чувствительным (лемнисковым) путям, поскольку даже после перерезки этих путей раздражение структур продолговатого мозга продолжало вызывать отчетливые изменения ЭЭГ.
Представления Дж. Моруцци и X. Мегуна об активирующей системе Р. ф. ствола мозга получили дальнейшее развитие и подтверждение в работах, выполненных в лабораториях многих стран мира. Были подтверждены основные выводы Дж. Моруцци и X. Мегуна и уточнена топография структур, относимых к восходящей активирующей ретикулярной системе мозгового ствола.
Электрофизиологические эксперименты, проведенные с помощью микроэлектродной техники, показали, что к одному и тому же нейрону Ретикулярной Формации могут сходиться импульсы от различных периферических источников и коры большого мозга. Взаимодействие между этими импульсами и обусловливает, по-видимому, многообразие наблюдаемых эффектов от внешних раздражений.
Работами Делля (Р. Dell) и его школы показано, что важная роль в поддержании активности Р. ф. принадлежит гуморальным факторам, в частности адреналину. Было установлено, что активирующее влияние адреналина на кору головного мозга осуществляется через Р. ф. среднего мозга и моста (рис. 2). После перерезки мозгового ствола кпереди от среднего мозга введение адреналина не вызывало больше «реакции пробуждения». Т. о., ростральные отделы мозгового ствола обладают повышенной чувствительностью к адреналину. Более того, исследованиями Фогта (М. Vogt, 4954) и П. К. Анохина (1956) в ростральных отделах мозгового ствола было выявлено наличие большого количества элементов, содержащих адреналин и норадреналин. И. П. Анохиной (1956) было обнаружено, что аминазин, обладающий способностью блокировать альфа-адренергические рецепторы, предотвращает развитие активации ЭЭГ при болевых раздражениях. Это дает основание предполагать, что активирующее влияние Р. ф. на кору головного мозга при болевых раздражениях осуществляется за счет вовлечения в активность а-адренергических рецепторов ростральных отделов мозгового ствола.
По каким путям осуществляется влияние ретикулярных механизмов на кору головного мозга? X. Мегун и Дж. Моруцци (1949) высказали предположение, что одним из путей, через к-рый Р. ф. оказывает влияние на кору головного мозга, является группа медиальных ядер таламуса (см.), составляющая так наз. неспецифическую проекционную таламокортикальную систему. К этой системе относятся (у кошки) nucleus reticularis, п. ventralis anterior, n. centralis medialis, n. lateralis, n. medialis, n. centrum medianum, n. parafascicularis и другие ядра интраламинарного комплекса (внутрипластинчатые ядра таламуса, Т.). При раздражении неспецифических ядер таламуса наблюдаются генерализованные изменения электрической активности коры головного мозга независимо от того, какое из ядер подвергается раздражению; ядра этой системы имеют между собой большое число связей, благодаря чему при возбуждении эта система реагирует как единая.
Изучению функциональных соотношений между восходящей активирующей системой Р. ф. мозгового ствола и неспецифической проекционной (диффузной) таламической системой посвящено большое число работ. Многие исследователи просто объединяют эти системы в одну, учитывая их тесное морфологическое единство. Однако Дж. Моруцци (1958) полагает, что для такого объединения нет достаточных оснований и неспецифическую таламическую систему целесообразнее относить к «сферам влияния» Р. ф., тем более что имеются экспериментальные данные, свидетельствующие о реципрокных взаимоотношениях между этими образованиями. Детальное изучение путей, связывающих Р. ф. мозгового ствола с неспецифическими ядрами таламуса, провел Пейпс (J. W. Papez, 1956). По его данным, таких путей три: ретикулярно-таламический, проходящий от Р. ф. продолговатого мозга к центромедиальному ядру таламуса, а также к заднемедиальному и парафасцикулярному ядрам. По латеральной стороне этого пути проходят чувствительные пути (лемнисковая система), от к-рых отходят коллатерали к ядрам покрышки. От ядер покрышки начинается тег-ментно-таламический путь, заканчивающийся в центральном срединном ядре (centrum medianum) и прилегающих к нему клетках. Самым латеральным по расположению является тектоталамический путь, оканчивающийся в пограничном ядре таламуса. Благодаря этим путям неспецифическая проекционная таламическая система становится связующим звеном между восходящей активирующей ретикулярной системой мозгового ствола и корой головного мозга.
Признание существования двух афферентных систем (специфической и неспецифической), имеющих разные формы, способы и сферы влияния, привело к необходимости изучения особенностей окончания волокон обеих этих систем в коре головного мозга. Было показано, что нервные волокна этих систем имеют окончания в коре, отличающиеся по форме и распределению в различных корковых слоях. Специфические афферентные волокна оканчиваются по преимуществу в 4-м слое коры, неспецифические волокна - во всех слоях коры. Специфические волокна оканчиваются главным образом на теле клетки, неспецифические - на ее дендритах. Аксодендритические окончания неспецифических волокон могут создавать изменение возбудимости корковых нейронов, облегчая или затрудняя синаптическую передачу (см. Синапс). Эти влияния диффузны и изменчивы. Аксосоматические окончания специфических волокон обеспечивают быстрые и локальные ответы. Взаимодействие обеих этих систем, по мнению Чанга (H. Т. Chang, 1952), обусловливает окончательную реакцию корковых нейронов.
Следует отметить, что если в 50-е и 60-е гг. 20 в. превалировал взгляд, согласно к-рому Р. ф. рассматривалась как диффузно организован-лая, оказывающая «неспецифическое» восходящее и нисходящее влияние, то в 70-е гг. это мнение стало пересматриваться.
Одним из первых исследователей, указавших на то, что активирующие влияния Р. ф. всегда имеют определенный биологический «знак», был П. К. Анохин (1958). Причиной радикального пересмотра взглядов явился прогресс как физиологических, так и морфологических методов исследования. К последним, в первую очередь, следует отнести совершенствование техники гистофлюоресцент-ного анализа, что позволило выявить в мозговом стволе нейроны, содержащие моноамины (норадреналин, серотонин и дофамин) и показать характер разветвления волокон этих нейронов. Отмечено, что норадрена-ловая и серотонинная системы содержат типичные ретикулярные нейроны. Эти данные заставили пересмотреть морфологическую догму, согласно к-рой все пути к коре головного мозга переключаются в таламусе. Было установлено, что мезенце--фалические серотонинергические и норадреналинергические нейроны восходят прямо от ядер Р. ф. в кору головного мозга. В связи с этим возник вопрос о нейрохимической основе корковой «реакции пробуждения».
Значительный вклад в физиологию Ретикулярной Формации внесло применение микроэлектродных методов исследования (см.). Было показано, что одним из важных механизмов активирующих ретикулоталамические и ретикуло-кортикальные влияния является подавление тормозных интернейронов, т. е. «торможение торможения».
Новые данные были получены в процессе изучения ретикуломотор-ной координации, т. е. в выявлении локальных групп клеток, специфически связанных с контролем определенных форм двигательной активности, в частности в идентификации нейронов, регулирующих движения глаз, нейронов, контролирующих механизмы позы и локомоции, и т. д.
Открытие в ряде структур Р. ф. высоких концентраций так наз. опиатных рецепторов, указывало на связь с организацией болевой чувствительности. К этим структурам относятся ядра срединного шва, также центральное серое вещество вокруг сильвиева водопровода и др.
Обнаружение функциональной специализации различных отделов Р. ф. привело к переоценке отношения к Р. ф. как к «неспецифической системе». В США был организован и проведен специальный международный симпозиум, материалы к-рого опубликованы в монографии «Пересмотр взглядов на ретикулярную формацию: специфическая функция неспецифической системы» (1980).
Однако надо отметить, что исследования, подытоживающие сведения о Р. ф., не отменяют взгляда на Р. ф. как на систему, работающую в функциональном единстве с анализаторными системами и оказывающую тонические влияния на ниже- и вышележащие отделы ц. н. с.
Нельзя не учитывать, что задолго до появления первых исследований, посвященных Р. ф., И. М. Сеченов показал, что в области мозгового ствола располагаются структуры, контролирующие деятельность спинного мозга. И. П. Павлов также придавал подкорковым образованиям большое значение в поддержании активности коры больших полушарий. Он говорил о «слепой силе» подкорки, о подкорке как «источнике энергии для коры». Установление важной роли ретикулярных механизмов в деятельности ц. н. с. является конкретизацией теоретических представлений этих гениальных русских ученых.
Патология ретикулярной формации
Нарушение функции Р. ф. развивается вследствие поражения ее ядер, локализующихся, гл. обр., в области продолговатого мозга, моста и среднего мозга, а также афферентных и эфферентных связей на различных уровнях.
Патология собственно интегративных функций Р. ф., как показали А. М. Вейн (1974), Илайессон (S. G. Eliasson, 1978), может проявляться в виде расстройств движений (см.), нарушений сознания (см.), сна (см.), вегетативной дисфункции.
Двигательные расстройства обусловлены нарушением фазического и тонического контроля поперечнополосатых мышц, к-рый в норме осуществляется посредством взаимодействия активирующих и тормозящих влияний Р. ф., передающихся на альфа- и гамма-мотонейроны спинного мозга (см.) через ретикулоспинальный и вестибулоспинальный пути.
Патология Р. ф. мозгового ствола, а также ее афферентных и эфферентных связей может сопровождаться как повышением мышечного тонуса и сухожильных рефлексов, так и их снижением. Мышечная гипертония и повышение сухожильных рефлексов возникают вследствие преобладания активирующих влияний ретикулярной формации на а- и у-мотонейроны спинного мозга, к-рое наблюдается при поражении гигантоклеточного ретикулярного ядра на уровне продолговатого мозга или моста, его афферентных связей с корой большого мозга и хвостатым ядром, а также при патологии эфферентных путей к мотонейронам спинного мозга.
Диффузное поражение Р. ф. на уровне мозгового ствола может приводить к резкому снижению мышечного тонуса и сухожильных рефлексов вследствие отсутствия активирующих влияний на а- и 7-мотонейроны спинного мозга.
Двигательные расстройства при патологии Р. ф. касаются не только поперечнополосатых мышц туловища и конечностей, но и мышц, иннервируемых черепно-мозговыми (черепными, Т.) нервами.
Другим клин, синдромом, наблюдаемым при патологии Р. ф., является расстройство сознания вплоть до появления комы. Кома (см.) характеризуется полной утратой сознания, отсутствием реакции на внешние раздражители, медленным синхронизированным ритмом ЭЭГ. В основе комы лежит блокада восходящей активирующей Р. ф., ответственной за процессы бодрствования, активации внимания на различные сенсорные стимулы. Функциональные или структурные нарушения восходящей активирующей системы на любом ее уровне, включая оральные отделы мозгового ствола, прозрачную перегородку, гипоталамус, таламус и таламокортикальные связи, могут приводить к расстройствам сознания. Коматозные состояния наиболее часто развиваются при патологии мозгового ствола и среднего мозга или при процессах, приводящих к их дислокации. Но возможен и другой механизм комы, при к-ром в ее основе лежит патология коры головного мозга и нарушение нисходящих влияний коры на Р.ф., в результате чего вторично изменяется функциональное состояние Р. ф.
Для поражения Р. ф. в области покрышки моста и среднего мозга характерен синдром псевдокомы, или акинетического мутизма. Синдром акинетического мутизма характеризуется потерей способности адекватно реагировать на внешние стимулы при сохранном сознании или слабо выраженном его нарушении. При этом у больного нарушается речь (см.), активные движения, он не запоминает события, происходящие в данный промежуток времени. Не изменяются зрачковые реакции, сухожильные и периостальные рефлексы. Сильные болевые и звуковые раздражения вызывают ответную двигательную реакцию. В основе синдрома лежит нарушение восходящей активирующей системы и ее связей с лимбическими структурами мозга (см. Лимбическая система), что приводит к отсутствию побуждений к действию, затруднению интеграции моторных функций, расстройству памяти (см.).
Частым симптомом поражения Р. ф. и ее связей является расстройство сна. Патология сна (см.) обусловлена либо нарушением функциональных реципрокных взаимоотношений между восходящей активирующей системой и гипногенными синхронизирующими зонами, ответственными за генерацию сна, либо дисфункцией самих гипногенных зон, расположенных гл. обр. в пределах лимбико-ретикулярного комплекса.
Расстройства сна могут быть в виде повышенной сонливости (гиперсомнии) и разнообразных нарушений ночного сна (инсомнии). Гиперсомнии могут быть обусловлены гипофункцией восходящей активирующей системы или гиперфункцией одной из систем механизмов регуляции сна. Гиперсомнические состояния, наблюдаемые при органическом поражении мезэнцефально-диэнцефальной области мозга, являются результатом нарушения функционирования активирующей системы Р. ф. Инсомнии, характеризующиеся бессонницей, трудностями засыпания, частыми просыпаниями, укорочением длительности ночного сна, могут быть обусловлены относительно повышенной функциональной активностью восходящей активирующей системы, а также нарушением функционирования отдельных сомноген-ных областей мозга, ответственных за генерацию фаз быстрого и медленного сна. Такие гипногенные зоны расположены в каудальном отделе мозгового ствола (так наз. синхронизирующая система Дж. Моруцци), в ядрах срединного шва, гипоталамусе. При их патологии наблюдается дезорганизация отдельных фаз сна. Нарушение циклической организации сна без грубых изменений отдельных фаз характерно для патологии интегрирующих аппаратов, регулирующих включение синхронизирующих и десинхронизирующих механизмов сна. К ним относятся отдельные структуры лимбико-ретикулярного комплекса (гипоталамус, зрительный бугор, базальные ядра конечного мозга), а также активирующая таламокортикальная система.
При нарушении функции Р. ф. мозгового ствола может наблюдаться синдром вегетативно-сосудистой дистонии. По данным А. М. Вейна и др. (1981), вегетативно-сосудистые расстройства отмечаются у большинства больных с патологией мозгового ствола. Вегетативные нарушения представлены кардиоваскулярными, вазомоторными и дыхательными расстройствами, к-рые могут иметь симпатико-адреналовую или парасимпатическую направленность (см. Нейроциркуляторная дистония). В основе вегетативных нарушений, возникающих при поражении Р. ф. мозгового ствола, лежит не только дисфункция специфических вегетативных центров (вазомоторного, дыхательного), но и нарушение целостной интегративной функции, необходимой для обеспечения целесообразного адаптивного поведения. Поэтому дыхательные, сердечно-сосудистые и вазомоторные расстройства, наблюдаемые при патологии Р. ф. мозгового ствола, сопровождаются изменениями мышечного тонуса, моторики и секреции внутренних органов, эндокринными нарушениями, изменениями настроения, снижением памяти. Имеется определенная зависимость клин, проявлений веге-тативно-сосудистой дистонии от области поражения Р. ф. мозгового ствола. При нарушении верхних отделов ствола вегетативные расстройства носят симпатическую направленность, могут сопровождаться легкими нейроэндокринными нарушениями. У больных с поражением каудальных отделов ствола выявляется парасимпатическая направленность тонуса вегетативной нервной системы, часто наблюдаются вестибулярные нарушения. Это обусловлено наличием связей Р. ф. с ядрами блуждающего нерва и вестибулярными ядрами.
Изучение физиологии и патофизиологии Р. ф. позволило значительно углубить представления о механизмах развития многих заболеваний нервной системы. Патология восходящей активирующей системы Р. ф. лежит в основе нарушений сознания и коматозных состояний (при острых нарушениях мозгового кровообращения, черепно-мозговых травмах, опухолях, энцефалитах, метаболических расстройствах). Блокада активирующих влияний на кору большого мозга может быть вызвана либо непосредственно самим очагом поражения, локализующимся в мозговом стволе, среднем мозге или гипоталамусе, либо отеком, приводящим к сдавлению, дислокации и вторичным метаболическим расстройствам в этой области.
При коматозных состояниях, обусловленных расстройством метаболизма (напр., при гипогликемии) или интоксикацией лекарственными препаратами (барбитураты, транквилизаторы, адренолитические средства), наблюдается прямое подавление нейронов активирующей ретикулярной системы или блокада адренергических рецепторов синапсов.
Повышение функциональной активности системы бодрствования, проявляющееся патологией сна в виде инсомний, характерно для неврозов. У больных с неврозами (см.) часто наблюдаются также синдром вегетативно-сосудистой дистонии, эмоциональные расстройства, характерные для дисфункции лимбико-ретикулярного комплекса.
Патология Р. ф. играет определенную роль и в развитии синдрома паркинсонизма (см.). Среди типичных морфологических изменений при этом заболевании часто обнаруживают гибель нейронов Р. ф., обеспечивающей состояние бодрствования. Повышенная сонливость и акинезия при паркинсонизме зависят не только от первичного поражения активирующей системы Р. ф., но и ее блокады за счет функционального усиления тормозных влияний хвостатого ядра на уровне ретику-локорковых связей.
Патология нисходящих влияний Р. ф. играет роль в формировании центральных параличей и парезов, экстрапирамидной ригидности, мио-клоний.
Уточнение функции Р. ф. и выяснение ее роли в развитии патол. нарушений стало возможным на основе широких экспериментальных и клинических исследований. Для изучения функции Р. ф. применяются метод вживления электродов с определением активности клеточных популяций, электроэнцефалографический анализ, морфологические исследования с применением электронной микроскопии (см.), методов гистохимии (см.) и биохимии (см.), в т. ч. изучение нейрохимии медиаторов (см.). В клин, практике широко используются полиграфические методы исследования, включающие одновременно электроэнцефалографию (см.), электроокулографию (см.), электромиографию (см.), электрокардиографию (см.), с помощью к-рых можно дифференцированно определять уровень поражения нервной системы, функциональное состояние восходящих и нисходящих систем Р. ф. и выявить особенности их реагирования на применение различных фармакологических средств.
Лечение
Имеется широкий арсенал фармакологических препаратов, воздействующих на функцию Р. ф. и ее связи с другими образованиями мозга. Селективным действием на активирующую ретикулярную систему обладают барбитураты, к-рые блокируют восходящие импульсы к коре головного мозга. Этот механизм лежит в основе их наркотического и противосудорожного эффектов. Прямое подавляющее действие на восходящую активирующую систему оказывают препараты брома, препараты фенотиазинового ряда (аминазин и др.), нек-рые транквилизаторы (хлордиазепоксид, диазепам, оксазепам, нитразепам), с чем связан их успокаивающий, противо-судорожный и легкий снотворный эффект. Восходящую ретикулярную систему активируют адренергические медиаторы (адреналин, норадреналин), их предшественник L-ДОФА, а также непрямые адреномиметики (кофеин, ниаламид, имизин, амитриптилин, фенамин, меридил, сиднокарб и др.). Эти средства используют в комплексной терапии больных в коматозном состоянии, при повышенной сонливости, депрессиях, астении. Холинергические синапсы ретикулярной формации блокируются центральными холиномиме-тиками (скополамин, амизил, мета-лизил), что приводит к снижению парасимпатических влияний Р. ф. мозгового ствола на внутренние органы. Уменьшение потока симпатических импульсов на периферию может быть достигнуто применением симпа-толитических средств (резерпин, метилдофа), к-рые нарушают образование катехоламинов и активируют тормозные структуры Р. ф. Существуют средства, оказывающие избирательное действие на серотонин-ергические структуры Р.ф. (L-триптофан, дизерил). Эти препараты используют в клинике с целью нормализации сна. Эффект торможения ретикулярных клеток каудального мозгового ствола такими препаратами, как лиорезал, мидокалам, диазепам, используется при лечении больных с повышением мышечного тонуса.
Коррекция синдромологических нарушений Р. ф. является частью комплексной терапии заболеваний нервной системы, в к-рой ведущее место должно отводиться этиологическому и патогенетическому лечению.
Библиография: Амунц В. В. Развитие ретикулярной формации ствола мозга в онтогенезе низшей обезьяны по сравнению с человеком, Арх. анат., гистол. и эмб-риол., т. 71, в. 7, с. 25, 1976, библиогр.; Анохин П. К. Значение ретикулярной формации для различных форм высшей нервной деятельности, Физиол. журн. СССР, т. 43, № 11, с. 1072,1957; о н ж е, Узловые вопросы теории функциональной системы, М., 1980; Б р о д а л А. Ретикулярная формация мозгового ствола, пер. с англ., М., 1960, библиогр.; Бейн А. М. Лекции по неврологии неспецифических систем мозга, М., 1974; Вейн А. М. и С о л о в ь е в а А. Д. Лимбико-ретикулярный комплекс и вегетативная регуляция, М., 1973, библиогр.; Вейн А. М., С о л о в ь е в а А. Д. и Колосова О. А. Вегетососудистая дистония, М., 1981, библиогр.; Д е-м и н H. Н., Коган А. Б. и М о и-сеева Н. И. Нейрофизиология и нейрохимия сна, Л., 1978; Жукова Г. П. Нейронное строение и межнейронные связи мозгового ствола и спинного мозга, М., 1977, библиогр.; Косицын Н. С. Микроструктура дендритов и аксодендри-тических связей в центральной нервной системе, М., 1976; М э г у н Г. Бодрствующий мозг, пер. с англ., М., 1961; Ретикулярная формация мозга, под ред. Г. Г. Джаспера и др., пер. с англ., М., 1962; Росси Д. Ф. и Ц а н к е т-т и А. Ретикулярная формация ствола мозга, пер. с англ., М., 1960, библиогр.; Свядощ А. М. Неврозы, М., 1982; Структура и функция ретикулярной формации и ее место в системе анализаторов, под ред. С. А. Саркисова, М., 1959; Handbook of clinical neurology, ed. by P. J. Vin-ken a. G. W. Bruyn, v. 1, Amsterdam a. o., 1975; Meessen H. u. Olszewski J. Cytoarchitektonischer Atlas des Rautenhirns des Kaninchens, Basel - N. Y., 1949; M o r u z z i G. a. M a- g o u n H. W. Brain stem reticular formation and activation of E E G, Electroen-ceph. clin. Neurophysiol., v. 1, p. 455, 1949; Neurological pathophysiology, ed. by S. G. Eliasson а. о., N. Y., 1978; О 1 s-zewski J. The cytoarchitecture of the human reticular formation, в кн.: Brain mechanisms and consciousness, ed. by E. D. Adrian a. o., p. 54, Oxford, 1954, bibliogr.; Purpura D. P., Me Murtry J. G. a. Maekawa K. Synaptic events in ventrolateral thalamic neurons during suppression of recruitfing responses by brain stem reticular stimulation, Brain Res., v. 1, p. 63, 1966; R a-mon у Cajal S. Histologie du sys-teme nerveux de l’homme et des vertebres, t. 1-2, Madrid, 1952-1955; The reticular formation revisited, ed. by J. A. Hobson а. М. A. B. Brazier, N. Y., 1980, bibliogr.
В. В. Амунц, В. Г. Скребицкий, В. Н. Шелихов; Л. О. Бадалян (невр.).